Lipoxygenase in a Giant Sulfur Bacterium: An Evolutionary Solution for Size and Complexity?
Georgy Kurakina
Pirogov Russian National Research Medical University, 117997 Moscow, Russia
Received April 17, 2023; Revised April 17, 2023; Accepted May 2, 2023
Discovery of Thiomargarita magnifica – an exceptionally large giant sulfur bacterium – urges us to pay additional attention to the giant sulfur bacteria and to revisit our recent bioinformatic finding of lipoxygenases in the representatives of the genus Beggiatoa. These close relatives of Thiomargarita magnifica meet the similar size requirements by forming multicellular structures. We hypothesize that their lipoxygenases are a part of the oxylipin signaling system that provides high level of cell-to-cell signaling complexity which, in turn, enables them to reach large sizes.
KEY WORDS: Beggiatoa, multicellularity, lipoxygenase, Thiomargarita magnificaDOI: 10.1134/S0006297923060111
INTRODUCTION
Oxylipins are oxidized derivatives of polyunsaturated fatty acids. They are involved in a wide range of cell-to-cell signaling processes in multicellular eukaryotes and function as:
- stress and injury mediators in animals, plants, and various groups of algae;
- pheromone-like compounds in fungi, brown and diatom algae;
- developmental signals in fungi and (partially) plants.
LIPOXYGENASE IN A GIANT SULFUR BACTERIUM
In our recent paper in Biochemistry (Moscow) [1], we presented the results of our bioinformatic study showing evolutionary associations of lipoxygenases with multicellularity (albeit in primitive forms) and host–microbe interactions (Fig. 1). This article aims to further discuss the first association – the link between lipoxygenases and multicellularity.
Fig. 1. Phylogenetic network of bacterial and eukaryotic lipoxygenases (from our previous article [1]) with ecophysiological mapping. Most lipoxygenase clusters are associated with multicellular organisms, and only a small sector consists of lipoxygenases of pathogenic (red arrows) and symbiotic (green arrow) bacteria. This fact suggests that bacteria need lipoxygenases either to be multicellular themselves or to interact with a multicellular host. Image credit: from Kurakin et al., 2020 [1] (amended).
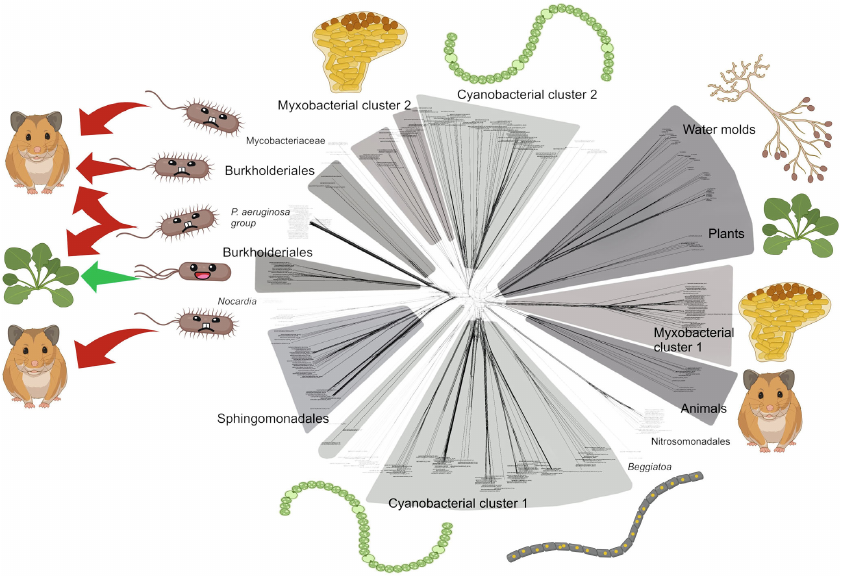
In the bacterial domain, presence of lipoxygenases and multicellularity co-occurred in cyanobacteria and myxobacteria. These two taxa are the best-known examples of bacterial multicellularity, and they form statistical picture of the higher prevalence of lipoxygenases in multicellular taxa, so they were included in the subsequent discussion.
However, there is a third bacterial taxon revealed to have LOXs in our previous paper. This is the genus Beggiatoa, which belongs to the group called “big sulfur bacteria” or “giant sulfur bacteria” [7, 8]. This name reflects their biochemistry: they are chemolithoautotrophs and derive their energy from oxidation of sulfide (S2–) to sulfate (SO42–) using oxygen or nitrate as oxidants. The name also reflects their unusually large size: Beggiatoa spp. form filaments from a few µm to 10 cm long, and the cell diameter ranges between ca. 1-200 µm. The filaments of Thioploca spp. are 2-5 cm long and 2.5-80 µm in diameter [9].
Representatives of these two genera (as well as some other genera of the giant sulfur bacteria) are multicellular. Their long filaments consist of several hundred cells [7], making them similar to the filamentous cyanobacteria. Despite this fact, we omitted them from our previous analysis [1] because LOXs were only detected in the genus Beggiatoa, which did not contribute significantly to the statistics of LOX occurrence at the order level and to the overall picture of statistical association between LOXs and multicellularity. However, it is worth noting that the presence of lipoxygenases in Beggiatoa was also confirmed by Chrisnasari et al. in their bioinformatic research [10].
But the recent discovery makes us look at this genus from a different perspective. In June 2022, a new giant sulfur bacterium was described in Science [11]. This bacterium, called Thiomargarita magnifica, can reach up to a centimeter long and could be visible to the naked eye. It is an all-time record for size in the bacterial world and has even been featured in the “2022 Breakthrough of the Year” special issue [12]. The fact that the genus Thiomargarita is one of the closest relatives of the genus Beggiatoa [7] suggests that any markers of the increased complexity among Beggiatoa species, including lipoxygenases, should be looked for. This is particularly important because, prior to the discovery of the genus Thiomargarita, Beggiatoa spp. were among the record holders for size: their cells can reach >100 µm [8]. With hundreds of such cells in a filament, they can reach up to several centimeters in length [7]. Thus, Beggiatoa genus is also mentioned in the context of challenging our understanding of size constraints in prokaryotes [8].
These size constraints arise from bioenergetics: ATP synthases, which supply cells with energy, can only be located on membranes, and in the most common bacteria, the only cell membrane is the cell envelope. The maximum cell size is therefore limited by the least surface-to-volume ratio, which still allows a cell to be sufficiently supplied with ATP.
On the other hand, giant sulfur bacteria live at the sulfide-oxygen or sulfide-nitrate interface and are forced to have sufficient length to bridge the sulfide pool and the oxidant pool to use them together in their metabolism. Moreover, they need to have sufficient cell volume. Most of the giant sulfur bacteria do this by evolving multicellular forms that allow them to form a large organism without exceeding the cell size limit. Thiomargarita magnifica is an interesting exception, which overcomes these constraints by increasing complexity of a cell using complex internal membrane network and nucleus-like organelles called pepins, where ATP synthases are located [11]. This increases the ATP-generating surface of membranes without building a multicellular organism.
Another Thiomargarita species – Thiomargarita namibiensis – also avoids being multicellular by using high nitrate storage capacity instead. They accumulate nitrate, when they are passively resuspended from the coastal sediment, and use it in further biochemical processes while buried in a sulfide-reach sediment. This strategy was metaphorically named “waiting strategy” [13] or “holding the microbe’s breath” [14].
The genera Beggiatoa, Thioploca, and Thiomargarita are close relatives in the family Beggiatoaceae, but they use different ways to fit in the oxidant-reductant interface. While Thiomargarita magnifica manages to do without being multicellular, both Thioploca and Beggiatoa form multicellular filaments. However, Thioploca species are also known for their more complex behavioral adaptation: they can actively move between the nitrate layer and sulfidic sediment, accumulating nitrate for its further use in the sulfidic environment [9, 15]. In contrast, Beggiatoa spp. depend on the simultaneous availability of both an oxidant (oxygen) and sulfide [9, 15]. Thus, multicellularity is almost the only strategy for Beggiatoa to bridge the two necessary substrates (Fig. 2).
Fig. 2. Beggiatoa, Thioploca, and Thiomargarita magnifica are close relatives that form exceptionally long structures to bridge two spatially distinct pools of sulfide (S2–, yellow) and oxygen or nitrate (O2/NO3–, blue). This creates selection pressure towards increase of the length (depicted by the arrows). But they use different strategies to overcome size constraints: T. magnifica enlarges the total ATP-generating surface by increasing complexity of a single cell, while Thioploca and Beggiatoa are multicellular. However, in the case of Thioploca, selection pressure is partially dampened by the ability of these bacteria to move up and down gliding along the mucus sheath. In contrast, Beggiatoa relies on bridging the pools of two compounds simultaneously and thus responds to the same level of selection pressure as Thiomargarita magnifica. We hypothesize that this evolutionary requirement is met by the increasing complexity of multicellular signaling, including oxylipin signaling, and Beggiatoa LOXs play essentially the same role as pepins and internal membranes in Thiomargarita magnifica.
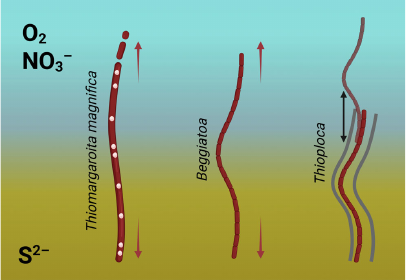
We suggest that this probably requires a higher level of cell-to-cell communication and integration. We hypothesize that this is the reason for the presence of lipoxygenases in Beggiatoa and their absence in Thioploca. These lipoxygenases may be the signature of oxylipin signaling in Beggiatoa species, which has developed in response to evolutionary demand for greater complexity.
This hypothesis fits well with our general assumption about the link between lipoxygenases, oxylipin signaling, and multicellularity. At present, this link is evident or suggested in animals, plants, fungi, water molds, dictyostelid slime molds, plasmodiophorids, cyanobacteria, and myxobacteria. We now add Beggiatoa spp. to this list. To validate this association, experimental studies of oxylipin signaling in the above bacterial taxa will be required.
Acknowledgments. Figures were created with BioRender.com. The author also acknowledges Alina Lebedeva (Tver State University, Teaching Assistant at the Department of Language Theory, Translation and French Philology) for literary editing this text.
Ethics declarations. The author declares no conflicts of interest in financial or any other sphere. This article does not contain description of studies with the involvement of humans or animal subjects performed by the author.
REFERENCES
1.Kurakin, G. F., Samoukina, A. M., and Potapova, N.
A. (2020) Bacterial and Protozoan lipoxygenases could be involved in
cell-to-cell signaling and immune response suppression, Biochemistry
(Moscow), 85, 1048-1063, doi: 10.1134/S0006297920090059.
2.Andreou, A. Z., Vanko, M., Bezakova, L., and
Feussner, I. (2008) Properties of a mini 9R-lipoxygenase from
Nostoc sp. PCC 7120 and its mutant forms, Phytochemistry,
69, 1832-1837, doi: 10.1016/j.phytochem.2008.03.002.
3.Lang, I., Göbel, C., Porzel, A., Heilmann, I.,
and Feussner, I. (2008) A lipoxygenase with linoleate diol synthase
activity from Nostoc sp. PCC 7120, Biochem. J.,
410, 347-357, doi: 10.1042/BJ20071277.
4.An, J. U., Hong, S. H., and Oh, D. K. (2018)
Regiospecificity of a novel bacterial lipoxygenase from Myxococcus
xanthus for polyunsaturated fatty acids, Biochim. Biophys. Acta
Mol. Cell Biol. Lipids, 1863, 823-833, doi:
10.1016/j.bbalip.2018.04.014.
5.Vance, R. E., Hong, S., Gronert, K., Serhan, C. N.,
and Mekalanos, J. J. (2004) The opportunistic pathogen Pseudomonas
aeruginosa carries a secretable arachidonate 15-lipoxygenase,
Proc. Natl. Acad. Sci. USA, 101, 2135-2139, doi:
10.1073/pnas.0307308101.
6.Morello, E., Pérez-Berezo, T., Boisseau, C.,
Baranek, T., Guillon, A., Bréa, D., Lanotte, P., Carpena, X.,
Pietrancosta, N., Hervé, V., Ramphal, R., Cenac, N., and
Si-Tahar, M. (2019) Pseudomonas aeruginosa lipoxygenase LoxA
contributes to lung infection by altering the host immune lipid
signaling, Front. Microbiol., 10, 1826, doi:
10.3389/fmicb.2019.01826.
7.Salman, V., Bailey, J. V., and Teske, A. (2013)
Phylogenetic and morphologic complexity of giant sulphur bacteria,
Antonie van Leeuwenhoek, 104, 169-186, doi:
10.1007/s10482-013-9952-y.
8.Jørgensen, B. (2010) Big sulfur bacteria,
ISME J., 4, 1083-1084, doi: 10.1038/ismej.2010.106.
9.Teske, A., and Nelson, D. C. (2006) in The
Prokaryotes (Dworkin, M., Falkow, S., Rosenberg, E., Schleifer,
K.-H., and Stackebrandt, E., eds) Springer, N. Y., pp. 784-810, doi:
10.1007/0-387-30746-X_27.
10.Chrisnasari, R., Hennebelle, M., Vincken, J. P.,
van Berkel, W. J., and Ewing, T. A. (2022) Bacterial lipoxygenases:
biochemical characteristics, molecular structure and potential
applications, Biotechnol. Adv., 61, 108046, doi:
10.1016/j.biotechadv.2022.108046.
11.Volland, J. M., Gonzalez-Rizzo, S., Gros, O.,
Tyml, T., Ivanova, N., Schulz, F., Goudeau, D., Elisabeth, N. H., Nath,
N., Udwary, D., Malmstrom, R. R., Guidi-Rontani, C., Bolte-Kluge, S.,
Davies, K. M., Jean, M. R., Mansot, J. L., Mouncey, N. J., Angert, E.
R., Woyke, T., and Date, S. V. (2022) A centimeter-long bacterium with
DNA contained in metabolically active, membrane-bound organelles,
Science, 376, 1453-1458, doi:
10.1126/science.abb3634.
12.Pennisi, E., Stokstad, E., Gibbons, A., Stone,
R., Cohen, J., Savitsky, Z., Voosen, P., Hutson, M., and Kaiser, J.
(2022) Runners-up, Science, 378, 1162-1167, doi:
10.1126/science.adg2799.
13.Schulz, H. N., and Riechmann, D. (2000)
Thiomargarita namibiensis: A giant with staying power [in
German], BioSpektrum, 6, 116-117.
14.Schulz, H. N. (2002) Thiomargarita
namibiensis: Giant microbe holding its breath, ASM News,
68, 122-127, doi: 10.1128/AEM.68.11.5746-5749.2002.
15.Kuenen, G. (1999) in Biology of the
Prokaryotes (Lengeler, J. W., Drews, G., and Schlegel, H. G., eds.)
Thieme Stuttgart, N. Y., pp. 234-260, doi:
10.1002/9781444313314.ch10.