Biochemical Characterization of Some Pyrazolopyrimidine-Based Inhibitors of Xanthine Oxidase
Hemlata Tamta, Sukirti Kalra, and Anup K. Mukhopadhyay*
Department of Biotechnology, National Institute of Pharmaceutical Education and Research, Sector 67, Phase X, S. A. S Nagar, Mohali, Punjab, India 160062; fax: 0091-172-214692; E-mail: akmukhopadhyay@niper.ac.in; Anup1niper@GMail.com* To whom correspondence should be addressed.
Received April 25, 2005; Revision received June 25, 2005
Inhibition of xanthine oxidase-catalyzed conversion of xanthine to uric acid by various pyrazolopyrimidine-based inhibitors (allopurinol derivatives) was evaluated and compared with the standard inhibitor allopurinol. Three compounds out of the seven compounds used in the study were found to be reasonably good inhibitors of xanthine oxidase (XO). 4-Amino-6-mercaptopyrazolo-3,4-d-pyrimidine was found to be the most potent inhibitor of XO (IC50 = 0.600 ± 0.009 µM). 4-Mercapto-1H-pyrazolo-3,4-d-pyrimidine (IC50 = 1.326 ± 0.013 µM) and 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine (IC50 = 1.564 ± 0.065 µM) also showed comparable inhibitory activity to that of allopurinol (IC50 = 0.776 ± 0.012 µM). All three compounds showed competitive type of inhibition with comparable Ki values. Induction of the electron transfer reaction catalyzed by XO in the presence of these compounds monitored as reduction of 2,6-dichlorophenolindophenol (DCPIP) revealed that electron transfer by 4-amino-6-mercaptopyrazolo-3,4-d-pyrimidine is comparable to that obtained by allopurinol or xanthine. However, 4-mercapto-1H-pyrazolo-3,4-d-pyrimidine and 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine did not show DCPIP reduction. On the other hand, enzymatic reduction of cytochrome c in the presence of the three compounds was found to be insignificant and much less in comparison to allopurinol and xanthine. Therefore, both 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine and 4-mercapto-1H-pyrazolo-3,4-d-pyrimidine displayed the inhibitory property and also did not produce XO-mediated reactive oxygen species (ROS). Since 4-mercapto-1H-pyrazolo-3,4-d-pyrimidine was found to have some toxicity, the effect of 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine on the enzymatic formation of uric acid and ROS was investigated and it was found that this compound was inhibiting enzymatic generation of both uric acid and ROS. It can be noted that the standard inhibitor, allopurinol, inhibits uric acid formation but produces ROS.
KEY WORDS: two-electron reduction, one-electron reduction, competitive inhibition, IC50, 4-amino-6-mercaptopyrazolo-3,4-d-pyrimidine, 4-mercapto-1H-pyrazolo-3,4-d-pyrimidine, 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine, allopurinol, xanthine, xanthine oxidaseDOI: 10.1134/S0006297906130086
Abbreviations: XO) xanthine oxidase; DCPIP) 2,6-dichlorophenolindophenol; ROS) reactive oxygen species.
Xanthine oxidase (XO) is a homodimer of catalytically independent
subunits with an approximate molecular mass of 150 kD each [1]. The enzyme catalyzes the conversion of
hypoxanthine to xanthine and xanthine to uric acid, which are the
terminal biochemical reactions of the purine degradation pathway [2]. Therefore, any defect in purine metabolism results
in an increase in the level of uric acid. This eventually leads to the
deposition of sodium hydrogen urate monohydrate crystals in joints. The
disease associated with this phenomenon is known as gout [3]. Moreover, increase in the level of XO also causes
renal stone formation and ischemic myocardium and reactive oxygen
species (ROS) induced diseases [4]. Allopurinol is
a suicide substrate of XO, which upon oxidation yields the product,
alloxanthine or oxypurinol (pyrazolo[3,4-d]pyrimidine-2,6-one). The
latter compound inactivates the enzyme by coordinating irreversibly to
the reduced form of XO, thus inhibiting formation of uric acid [5]. The mechanism of inhibition of xanthine oxidase
involves enzymatic conversion of allopurinol into oxypurinol, which
binds stoichiometrically to the reduced Mo(IV) [6].
Although the inhibitor binds very tightly to the enzyme, the inhibition
is time dependent [6, 7].
Furthermore, it is necessary to maintain an effective concentration of
the inhibitor in the organ because the inhibitor dissociates from the
enzyme by spontaneous oxidation of molybdenum Mo(IV) to Mo(VI), with a
half-life of 300 min at 25°C, with concomitant recovery of enzyme
activity [6]. Apart from allopurinol, several
pteridines [8], thiazoles [9],
phenyl pyrazoles [10], aryl triazoles [11], and flavonoids [12, 13] are also reported to be inhibitors of xanthine
oxidase. In the present study, we have systematically selected some
pyrazolopyrimidine-based compounds (allopurinol derivatives) and
investigated detailed inhibition mechanisms and induction of any
enzymatic electron transfer reaction initiated by these compounds in an
attempt to find some pyrazolopyrimidine- or allopurinol-based
inhibitors with unique characteristics (i.e., inhibitor of both
XO-mediated uric acid and ROS generation). Recently, we systematically
screened and investigated the inhibition mechanism and induction of
enzymatic electron transfer reaction of purine based compounds and
found that one of the representative compounds of this group,
6-(N-benzoylamino)purine, is a novel and potent inhibitor of XO
[14] with the abovementioned unique
characteristics.
MATERIALS AND METHODS
Chemicals. Xanthine oxidase, xanthine, o-dianisidine, cytochrome c, and allopurinol were purchased from Sigma (USA). 4-Mercapto-1H-pyrazolo-3,4-d-pyrimidine, 4-amino-6-mercaptopyrazolo-3,4-d-pyrimidine, and 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine were obtained from Lancaster (UK). All other allopurinol-based compounds listed in Table 1 were purchased from Lancaster. Hydrogen peroxide (30% v/v) was obtained from Merck (India).
XO activity assay. The enzyme activity was monitored spectrophotometrically by measuring uric acid formation at 293 nm with saturated concentration of xanthine (20 µM) as the substrate (if not mentioned otherwise for a specific experiment) in 1 ml of 50 mM phosphate buffer, pH 7.5, at 25°C [15]. In all cases the reaction was started by the addition of 12 nM of XO.
XO inhibition. Inhibition of XO-catalyzed xanthine to uric acid formation reaction by various purine-based inhibitors was measured by observing the decrease in the uric acid formation at 293 nm. The assay mixture contained both the substrate and the inhibitor in order to have equal competition of the substrate and inhibitor for the enzyme active site.
Unit definition. One unit of xanthine oxidase is defined as the amount of the enzyme that produces 1 nmol of uric acid in 1 ml of assay volume per min at 25°C or it is monitored as the amount of the enzyme that is responsible for a change in absorbance of 0.011 at 293 nm in 1 ml assay solution in 1 min at 25°C. Concentration of the enzyme was calculated using the molar extinction coefficient of XO (epsilon450 nm= 36 mM-1*cm-1) [16]. The pH and the concentration of XO used for all our assays were 7.5 and 2.25 units/ml, respectively, for all the assays.
Superoxide estimation. XO-mediated generation of superoxide was measured as an increase in the absorbance at 550 nm due to the reduction of cytochrome c (Deltaepsilon_= 21,000 M-1*cm-1) [14, 17].
Hydrogen peroxide estimation. The amount of hydrogen peroxide was determined using a coupled assay containing o-dianisidine and horseradish peroxidase. The reaction was terminated by adding 25% of sulfuric acid (H2SO4) and the absorbance was measured at 510 nm [14, 18].
Software and standard deviation. All our kinetic and IC50 data were fitted using the equation for nonlinear regression analysis. Grafit software (Erithacus Software Limited, UK), version 4.0, written by Dr. Robin J. Leatherbarrow, was used for this purpose. Each experimental value is the mean of three individual experiments. Values are expressed as mean ± SEM. Statistical significance was set as p < 0.05.
The Lineweaver-Burk equation was used for the calculation of Ki (equilibrium inhibition constant) for competitive inhibition:
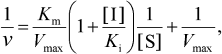
RESULTS AND DISCUSSION
Search for allopurinol-based inhibitors of XO. Time dependent spectra (from 200 to 400 nm) of all the allopurinol-based compounds were taken and the inhibition effect of these compounds on the XO-catalyzed xanthine to uric acid reaction was monitored (for the reaction chemistry, see Scheme). The structures of all the compounds used are shown in Table 1. All these compounds were also incubated with XO and their specificity as substrate of XO was monitored. These compounds were not found to be XO substrates, as they did not show any change in absorbance with time at their corresponding lambdamax. Subsequently, the inhibitory effect of these compounds on the enzymatic uric acid formation from the substrate xanthine was evaluated in the presence of various concentrations of these inhibitors. 4-Amino-6-hydroxypyrazolo-3,4-d-pyrimidine (IC50 = 1.564 ± 0.065 µM), 4-mercapto-1H-pyrazolo-3,4-d-pyrimidine (IC50 = 1.326 ± 0.013 µM), and 4-amino-6-mercaptopyrazolo-3,4-d-pyrimidine (IC50 = 0.600 ± 0.009 µM) showed activity comparable to allopurinol (IC50 = 0.776 ± 0.012 µM) (Table 1). None of the other compounds showed substantial inhibition of xanthine oxidase-mediated conversion of xanthine to uric acid.
Table 1. Structure and IC50
values of allopurinol and pyrazolopyrimidine-based inhibitors of
xanthine oxidase (all the IC50 values were calculated from
the percentage inhibition of the xanthine oxidase-mediated conversion
of xanthine (10 µM) to uric acid)
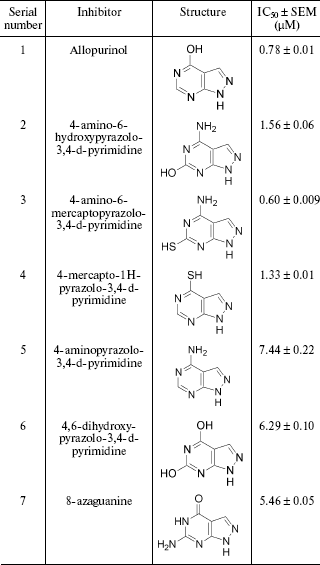
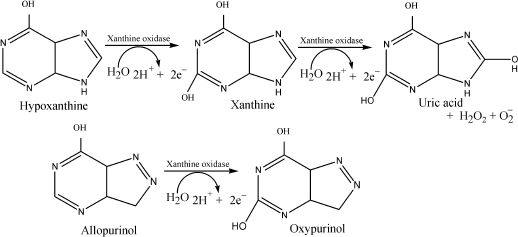
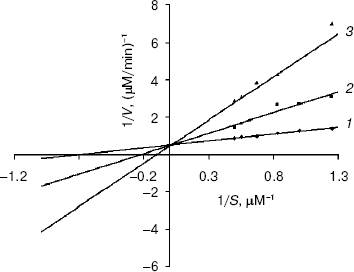
Table 2. Ki values of xanthine oxidase inhibitors, 4-mercapto-1H-pyrazolo-3,4-d-pyramidine and 4-amino-6-mercaptopyrazolo-3,4-d-pyramidine (enzyme concentration used was 3.25 units/ml or 12 nM; the xanthine concentrations taken were 0.8, 1.0, 1.2, 1.5, and 2.0 µM)Fig. 1. Lineweaver-Burk or double reciprocal plot for the inhibition of xanthine oxidase by 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine. The concentration of the enzyme was 3.25 units/ml or 1 nM. The substrate concentrations taken were 0.8, 1.0, 1.2, 1.5, and 2.0 µM. Each data point is the mean of experimental values from three individual experiments: 1) no inhibitor; 2) inhibitor concentration is 0.6 µM; 3) inhibitor concentration is 1.0 µM.

Note: The reaction rate was saturated with 10 µM xanthine; 10 and 20 µM xanthine exhibited identical reaction rate.
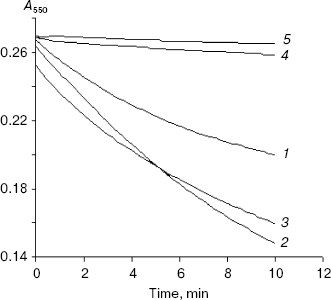
Status of the XO-mediated electron transfer reaction in the presence of inhibitors--xanthine and allopurinol. Initiation of the electron transfer reaction of XO in the presence of these allopurinol-based inhibitors was measured as the reduction of 2,6-dichlorophenolindophenol (DCPIP) (as electron acceptor redox dye) at 605 nm. 4-Amino-6-mercaptopyrazolo-3,4-d-pyrimidine showed maximum reduction of DCPIP. The result therefore reveals that the inhibitor binds to the enzyme active site and transfers electrons to the molybdenum center, which is then transferred to the electron acceptor dye, DCPIP (a two electron acceptor from the Mo site of XO). Figure 2 shows the comparison of DCPIP reduction kinetics by XO in the presence of xanthine, allopurinol, 4-amino-6-mercaptopyrazolo-3,4-d-pyrimidine, 4-mercapto-1H-pyrazolo-3,4-d-pyrimidine, and 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine. The enzymatic reduction progress curve of DCPIP in the presence of 4-amino-6-mercaptopyrazolo-3,4-d-pyrimidine is found to be comparable to xanthine and allopurinol. However, DCPIP reduction was not observed in the presence of 4-mercapto-1H-pyrazolo-3,4-d-pyrimidine and 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine. All the control experiments where these inhibitors and DCPIP were mixed together in the absence of xanthine oxidase did not show any appreciable DCPIP reduction, suggesting thereby that non-enzymatic reduction was absent.
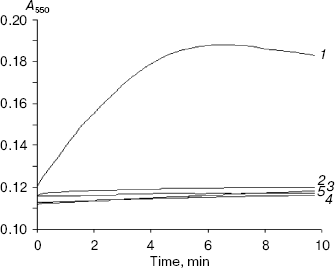
Status of XO-mediated superoxide formation in the presence of inhibitors--xanthine and allopurinol. XO-catalyzed superoxide production using xanthine, allopurinol, 4-amino-6-mercaptopyrazolo-3,4-d-pyrimidine, 4-mercapto-1H-pyrazolo-3,4-d-pyrimidine, and 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine as substrates was determined by monitoring the reduction of cytochrome c (a one electron acceptor) at 550 nm (Fig. 3). None of these inhibitors induced any enzymatic superoxide formation, unlike xanthine. Allopurinol induced a very minute amount of superoxide formation, which is insignificant as compared to xanthine. It is however important to mention here that the standard inhibitor (a suicide substrate) allopurinol reduces DCPIP (a two-electron acceptor) at a considerable rate and also the total extent of reduction is comparable to that from xanthine as the substrate. But interestingly, allopurinol as the substrate of XO did not cause any enzymatic reduction of cytochrome c (a one-electron acceptor). The result therefore suggests that allopurinol donates two electrons (DCPIP is a two-electron acceptor) to xanthine oxidase during its conversion to the suicide enzyme inhibitor oxypurinol but it does not reduce the enzyme via one-electron transfer (cytochrome c is a one-electron acceptor).Fig. 2. Kinetics of xanthine oxidase-mediated DCPIP reduction in the presence of xanthine (1), allopurinol (2), and the following allopurinol-based inhibitors: 3) 4-amino-6-mercapto-3,4-d-pyrazolopyrimidine; 4) 4-mercapto-1H-3,4-d-pyrazolopyrimidine; 5) 4-amino-6-hydroxy-3,4-d-pyrazolopyrimidine. The concentration of each compound used was 10 µM. DCPIP concentration was fixed at 15 µM.
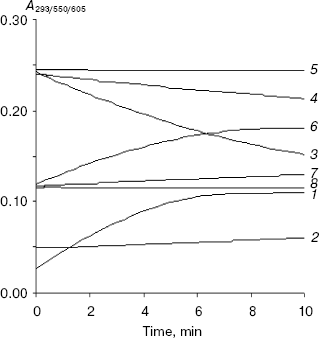
Inhibition of XO-mediated uric acid and ROS formation by 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine. Of the three compounds, since 4-amino-6-mercaptopyrazolo-3,4-d-pyrimidine induced the enzymatic reduction of DCPIP and 4-mercapto-1H-pyrazolo-3,4-d-pyrimidine displayed some toxicity in animal studies, we selected only 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine for studying the inhibition of XO-mediated formation of uric acid and ROS (see Scheme for the reaction). Figure 4 shows very clearly that this compound inhibited uric acid formation (curves 1 and 2), DCPIP reduction (curves 3 and 4), and superoxide formation (as monitored by cytochrome c reduction) (curves 6 and 7) very effectively. Furthermore, it is seen from Fig. 4 (curves 5 and 8) that 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine did not induce the xanthine oxidase-mediated reduction of DCPIP and cytochrome c and therefore does not produce enzyme-mediated reactive oxygen species on its own. Thus, the results suggest that this compound may be taken for further evaluation of in vivo studies.Fig. 3. Kinetics of xanthine oxidase-mediated superoxide formation in the presence of xanthine (1), allopurinol (2), and the following allopurinol-based inhibitors: 3) 4-amino-6-mercapto-3,4-d-pyrazolopyrimidine; 4) 4-mercapto-1H-3,4-d-pyrazolopyrimidine; 5) 4-amino-6-hydroxy-3,4-d-pyrazolopyrimidine. The concentration of each compound used was 10 µM. Cytochrome c concentration was fixed at 15 µM.
In this work, we have systematically investigated the inhibition mechanism and electron transfer properties of a few allopurinol or pyrazolopyrimidine-based compounds. Out of seven compounds (allopurinol derivatives) tested, 4-mercapto-1H-pyrazolo-3,4-d-pyrimidine, 4-amino-6-mercaptopyrazolo-3,4-d-pyrimidine, and 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine showed inhibitory activity comparable to that of allopurinol. These three compounds were found to be competitive inhibitors of XO. It was already shown that allopurinol reduced DCPIP by a XO-mediated two-electron transfer mechanism but did not generate superoxide (as monitored by cytochrome c reduction assay), which is a one-electron product. When allopurinol was used as the substrate of XO, its monohydroxylated product oxypurinol, was not detected optically (optically silent) and therefore, allopurinol can be called a silent suicide substrate. Among the allopurinol-based inhibitors, 4-amino-6-mercaptopyrazolo-3,4-d-pyrimidine displayed enzyme-mediated DCPIP reduction but did not induce any enzymatic superoxide formation and, therefore, this compound released only two electrons on interaction with XO. On the other hand, 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine and 4-mercapto-1H-pyrazolo-3,4-d-pyrimidine neither reduced DCPIP nor induced the formation of enzymatic superoxide, indicating thereby that these inhibitors did not release enzyme-mediated two or one electron species. It has been shown here that the mechanism of inhibition of XO by 4-mercapto-1H-pyrazolo-3,4-d-pyrimidine and 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine is different from that of allopurinol, while 4-amino-6-mercaptopyrazolo-3,4-d-pyrimidine acts very much like allopurinol in terms of enzyme-mediated electron transfer activity. However, 4-amino-6-mercaptopyrazolo-3,4-d-pyrimidine is a purely competitive inhibitor of XO, whereas allopurinol is a known suicide substrate of XO. 4-Amino-6-mercaptopyrazolo-3,4-d-pyrimidine is reported to be a toxic compound (as stated by Lancaster catalog) probably due to the presence of sulfur. Therefore, the inhibition (uric acid) and antiradical property (ROS) of only 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine was investigated in this work and we found that it effectively inhibited the enzymatic formation of uric acid and ROS as well.Fig. 4. Inhibition of enzymatic uric acid and reactive oxygen species formation from xanthine by 4-amino-6-hydroxy-3,4-d-pyrazolopyrimidine: 1) uric acid from 10 µM xanthine (as substrate); 2) uric acid from 10 µM xanthine in presence of 10 µM 4-amino-6-hydroxy-3,4-d-pyrazolopyrimidine (as inhibitor) (curves 1 and 2 were monitored at 293 nm); 3) enzymatic DCPIP reduction using 10 µM xanthine (as substrate); 4) enzymatic DCPIP reduction from xanthine (as substrate) in the presence of 10 µM 4-amino-6-hydroxy-3,4-d-pyrazolopyrimidine (as inhibitor); 5) enzymatic DCPIP reduction from 10 µM 4-amino-6-hydroxy-3,4-d-pyrazolopyrimidine (as substrate) (curves 3, 4, and 5 were monitored at 605 nm); 6) enzymatic cytochrome c reduction from 10 µM xanthine (as substrate); 7) cytochrome c reduction from 10 µM xanthine (as substrate) in the presence of 10 µM 4-amino-6-hydroxy-3,4-d-pyrazolopyrimidine (as inhibitor); 8) cytochrome c reduction from 10 µM 4-amino-6-hydroxy-3,4-d-pyrazolopyrimidine (as substrate) (curves 6, 7, and 8 were monitored at 550 nm). Assay conditions (pH and temperature) and the assay protocol are given in “Materials and Methods”.
There is overwhelming acceptance that the level of XO in the serum is significantly increased in several pathological states and that ROS generated in the enzymatic reaction are involved in the oxidative damage of different macromolecules. Thus, inhibition of this enzymatic pathway by some compounds that have both antiradical and uric acid inhibition properties could have therapeutic interest. Of the three compounds tested in our present work, only 4-amino-6-hydroxypyrazolo-3,4-d-pyrimidine displayed this property beyond any doubt.
Recently in our laboratory, we found a novel and potent purine-based inhibitor [14], 6-(N-benzoylamino)purine, which also displayed both uric acid inhibitory as well as antiradical property. At present, we are focusing on the study of the pharmacological and toxicity profile of the two compounds reported above.
We also thank the Council of Scientific and Industrial Research (CSIR, New Delhi, Government of India, grant No. 01(1823)/02/EMR-II) for funding of the project.
REFERENCES
1.Hille, R., and Nishino, T. (1995) FASEB J.,
9, 995-1003.
2.Krenitsky, T. A., Spector, T., and Hall, W. W.
(1986) Arch. Biochem. Biophys., 247, 108-119.
3.Rodnan, G. P. (1982) Bull. Rheum. Dis.,
32, 43-53.
4.McCord, J. M. (1985) N. Engl. J. Med.,
312, 159-163.
5.Nakamura, M. (1991) J. Biochem. (Tokyo),
110, 450-456.
6.Massey, V., Komai, H., Palmer, G., and Elion, G. B.
(1970) J. Biol. Chem., 245, 2837-2844.
7.Spector, T., and Johns, D. G. (1970) J. Biol.
Chem., 245, 5079-5085.
8.Oettl, K., and Reibnegger, G. (1999) Biochim.
Biophys. Acta, 1430, 387-395.
9.Okamoto, K., Eger, B. T., Nishino, T., Kondo, S.,
and Pai, E. F. (2003) J. Biol. Chem., 278, 1848-1855.
10.Ishibuchi, S., Morimoto, H., Oe, T., Ikebe, T.,
Inoue, H., Fukunari, A., Kamezawa, M., Yamada, I., and Naka, Y. (2001)
Bioorg. Med. Chem. Lett., 11, 879-882.
11.Okamoto, K., Matsumoto, K., Hille, R., Eger, B.
T., Pai, E. F., and Nishino, T. (2004) Proc. Natl. Acad. Sci.
USA, 101, 7931-7936.
12.Lin, C. M., Chen, C. S., Chen, C. T., Liang, Y.
C., and Lin, J. K. (2002) Biochem. Biophys. Res. Commun.,
294, 167-172.
13.Van Hoorn, D. E., Nijveldt, R. J., van Leeuwen,
P. A., Hofman, Z., M'Rabet, L., De Bont, D. B., and van Norren, K.
(2002) Eur. J. Pharmacol., 451, 111-118.
14.Tamta, H., Thilagavathi, R., Chakraborti, A. K.,
and Mukhopadhyay, A. K. (2005) J. Enzyme Inhibition Med. Chem.,
20, 317-324.
15.Escribano, J., Gracia-Canovas, F., and
Gracia-Carmona, F. (1988) Biochem. J., 254, 829-833.
16.Godner, B. L. J., Doel, J. J., Goult, T. A.,
Eisenthal, R., and Harrison, R. (2001) Biochem. J., 358,
325-333.
17.Hodges, G. R., Young, M. J., Paul, T., and
Ingold, K. U. (2000) Free Rad. Biol. Med., 29,
434-441.
18.Liochev, S. I., and Fridovich, I. (2002) J.
Biol. Chem., 277, 34674-34678.